Pentru a putea vorbi într-o manieră inteligibilă despre hemoglobină (Hb), este util să tratăm mai întâi mioglobina (Mb) care este foarte asemănătoare cu hemoglobina, dar este mult mai simplă. Între hemoglobina și mioglobina există relații stricte de rudenie: ambele sunt proteine conjugate și grupul protetic (partea non-proteică) este grupul heme .
Myoglobina este o proteină globulară formată dintr-un singur lanț de aproximativ o sută cincizeci de aminoacizi (depinde de organism) și greutatea sa moleculară este de aproximativ 18 Kd.
Așa cum sa spus, este înzestrat cu un grup de heme care este inserat într-o porțiune hidrofobă (sau lipofilă) a proteinei, constând din pliuri legate de structurile α-helix ale proteinelor fibroase.
Myoglobina constă în principal din segmente a-helices, prezente în număr de opt și constă aproape exclusiv din reziduuri nepolare (leucină, valină, metionină și fenilalanină), în timp ce reziduurile polar sunt practic absente (acid aspartic, acid glutamic, lizină și arginină); singurele reziduuri polare sunt două histidine, care joacă un rol fundamental în atacul oxigenului la grupul heme.
Grupul heme este un grup cromofor (absoarbe în vizibil) și este grupul funcțional de mioglobină.
Vezi deasemenea: hemoglobină glicozită - hemoglobină în urină
Un pic de chimie
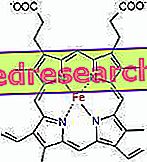
Legătura dintre protoporfirină și fier este o legătură tipică a așa-numitelor compuși de coordonare care sunt compuși chimici în care un atom central (sau ion) formează legături cu alte specii chimice într-un număr mai mare decât numărul său de oxidare (încărcătură electrică). În cazul heme, aceste legături sunt reversibile și slabe.
Numărul de coordonate (numărul de legături de coordonare) al fierului este de șase: în jurul fierului pot exista șase molecule care împart electronii de legătură.
Pentru a forma un compus de coordonare, este nevoie de două orbite cu orientarea corectă: una capabilă să "cumpere" electroni, iar cealaltă să le poată dona
În hem, fierul formează patru legături plane cu cei patru atomi de azot localizați în centrul inelului proto-porfirin și o a cincea legătură cu un azot al histidinei proximale; fierul are a șasea legătură de coordonare liberă și se poate lega de oxigen.
Atunci când fierul este sub forma unui ion liber, orbitele lui de tip d au toate aceeași energie; în mioglobină, ionul de fier este legat de protoporfirină și histidină: aceste specii perturba magnetic orbitalul d de fier; mărimea perturbării va fi diferită pentru diferitele orbite d în funcție de orientarea lor spațială și cea a speciilor perturbatoare. Întrucât energia totală a orbitalilor trebuie să fie constantă, perturbarea determină o separare energetică între diferitele orbite: energia obținută de unele orbite, echivalează cu energia pierdută de celelalte.
Dacă separarea care are loc între orbitale nu este foarte mare, este preferabil un aranjament electronic de înaltă înțepătură: electronii de legare încearcă să aranjeze rotația paralelă în mai multe sub-nivele posibile (multiplicitatea maximă); dacă, pe de altă parte, perturbația este foarte puternică și există o mare separare între orbitali, poate fi mai convenabil să se cupleze electronii de legare în orbitalele de energie inferioare (rotația joasă).
Atunci când fierul se leagă de oxigen, molecula are un aranjament scăzut, în timp ce atunci când fierul are cea de-a șasea legătură de coordonare liberă, molecula dispune de o rotire înaltă.
Datorită acestei diferențe în rotație, printr-o analiză spectrală a mioglobinei, suntem capabili să înțelegem dacă oxigenul este legat (MbO2) sau nu (Mb).
Myoglobina este o proteină tipică pentru mușchi (dar nu se găsește numai în mușchi).
Myoglobina este extrasă din balena spermei în care este prezentă în cantități mari și apoi purificată.
Cetaceenii au o respirație ca aceea a ființelor umane: având plămânii să absoarbă aerul prin procesul respirator; balena de spermă trebuie să aducă cât mai mult oxigen posibil în mușchii care sunt capabili să acumuleze oxigen prin legarea lui la mioglobina prezentă în ele; oxigenul este apoi eliberat încet când cetaceanul este scufundat, deoarece metabolismul lui necesită oxigen: cu cât este mai mare cantitatea de oxigen pe care balena spermatozoizilor este capabilă să o absoarbă, cu atât este mai mare oxigenul disponibil în timpul scufundării.

Pentru a face carnea mai mult sau mai puțin roșie este conținutul de hemoproteine (este hema care face carnea roșie).
Hemoglobina are multe analogii structurale cu mioglobina si este capabila de a lega oxigenul molecular in mod reversibil; dar, în timp ce mioglobina se limitează la mușchii și țesuturile periferice în general, hemoglobina se găsește în eritrocite sau celule roșii (ele sunt celule pseudo-celule, adică nu sunt celule reale) care reprezintă 40% din sânge.
Hemoglobina este un tetrametru, adică este alcătuit din patru lanțuri polipeptidice, fiecare având o grupare hemă și identice două câte două (la o ființă umană există două lanțuri alfa și două lanțuri beta).
Funcția principală a hemoglobinei este transportul de oxigen ; o altă funcție a sângelui în care este implicată hemoglobina este transportul substanțelor în țesuturi.
În calea plămânilor (bogată în oxigen) către țesuturi, hemoglobina transportă oxigen (în același timp, celelalte substanțe ajung în țesuturi) în timp ce în calea inversă, aduce împreună cu acesta deșeurile colectate din țesuturi, în special dioxidul de carbon produs în metabolism.
În dezvoltarea unei ființe umane există gene care sunt exprimate numai pentru o anumită perioadă de timp; din acest motiv avem hemoglobine diferite: fetale, embrionare, ale omului adult.
Lanțurile care alcătuiesc aceste hemoglobine diferite au structuri diferite, dar cu similitudini, funcția pe care o efectuează este mai mult sau mai puțin aceeași.
O explicație a prezenței mai multor lanțuri diferite este următoarea: în timpul procesului evolutiv de organisme, hemoglobina a evoluat și ea, specializată în transportul de oxigen din zonele bogate în zone deficitare. La începutul lanțului evolutiv, hemoglobina a transportat oxigenul în organisme mici; în cursul evoluției, organismele au atins dimensiuni mai mari, prin urmare hemoglobina sa schimbat pentru a putea transporta oxigen în zone mai îndepărtate de punctul în care era bogat; pentru a face acest lucru, noi structuri ale lanțurilor care alcătuiesc hemoglobina au fost codificate în cursul procesului evolutiv.
Myoglobinul leagă oxigenul chiar și la presiuni modeste; în țesuturile periferice există o presiune (PO2) de aproximativ 30 mmHg: mioglobina la această presiune nu eliberează oxigen, deci ar fi ineficient ca un transportor de oxigen. Hemoglobina, pe de altă parte, are un comportament mai elastic: se leagă de oxigen la presiuni mari și îl eliberează atunci când presiunea scade.
Când o proteină este activă din punct de vedere funcțional, ea își poate schimba puțin forma; de exemplu, mioglobina oxigenată are o formă diferită de cea a myoglobinei ne-oxigenate și această mutație nu afectează cele vecine.
Discursul este diferit în cazul proteinelor asociate, cum ar fi hemoglobina: când un lanț este oxigenat, acesta este indus să-și schimbe forma, dar această modificare este tridimensională și prin urmare celelalte lanțuri ale tetrametrului sunt, de asemenea, afectate. Faptul că lanțurile sunt asociate reciproc, conduce pe cineva să creadă că modificarea unuia afectează ceilalți vecini, chiar dacă într-o măsură diferită; când un lanț este oxigenat, celelalte lanțuri ale tetrameterului iau o "atitudine mai puțin ostilă" față de oxigen: dificultatea cu care un oxigenat al lanțului scade, pe măsură ce lanțurile apropiate acestuia oxigenază la rândul său. Același discurs este valabil pentru dezoxigenare.
Structura cuaternară a deoxihemoglobinei ia numele formei T (tesa), în timp ce cea a oxihemoglobinei este denumită forma R (eliberată); în starea tensionată există o serie de interacțiuni electrostatice destul de puternice între aminoacizii și aminoacizii de bază care duc la o structură rigidă de deoxihemoglobină (de aceea forma "tensionată"), în timp ce atunci când oxigenul este legat, magnitudinea acestora interacțiunile scad (de aceea "forma eliberată"). Mai mult decât atât, în absența oxigenului, încărcarea histidină (vezi structura) este stabilizată prin încărcarea opusă a acidului aspartic, în timp ce, în prezența oxigenului, există o tendință din partea proteinei de a pierde un proton; toate acestea înseamnă că hemoglobina oxigenată este un acid mai puternic decât efectul hemoglobinei dezoxigenate: bohr .
În funcție de pH, grupul heme se leagă mai mult sau mai puțin ușor de oxigen: într-un mediu acid, hemoglobina eliberează oxigenul mai ușor (forma tensionată este stabilă), în timp ce într-un mediu de bază, legătura cu oxigen este mai puternic.

Fiecare hemoglobină eliberează 0, 7 protoni pe mol de oxigen de intrare (O2).
Efectul Bohr permite hemoglobinei să își îmbunătățească capacitatea de a transporta oxigen.
Hemoglobina care face călătoria de la plămâni la țesuturi trebuie să se echilibreze în funcție de presiune, pH și temperatură.
Să vedem efectul temperaturii .
Temperatura în alveolele pulmonare este cu aproximativ 1-1, 5 ° C mai mică decât temperatura exterioară, în timp ce în mușchi temperatura este de aproximativ 36, 5-37 ° C; pe măsură ce crește temperatura, factorul de saturație scade (la aceeași presiune): acest lucru se întâmplă deoarece crește energia cinetică și este favorizată disocierea.

Există și alți factori care pot afecta capacitatea hemoglobinei de a se lega de oxigen, una dintre acestea fiind concentrația de 2, 3 bisfosfoglicerat .
2, 3 bisfosfogliceratul este un metabolic prezent în eritrocite la o concentrație de 4-5 mM (în nici o altă parte a corpului nu este prezent într-o concentrație atât de mare).
La pH fiziologic, 2, 3-bisfosfogliceratul este deprotonat și are cinci sarcini negative asupra acestuia; este legată între cele două lanțuri beta ale hemoglobinei, deoarece aceste lanțuri au o concentrație ridicată de încărcături pozitive. Interacțiunile electrostatice dintre lanțurile beta și 2, 3-bisfosfogliceratul dau o anumită rigiditate sistemului: se obține o structură tensionată care are o mică afinitate pentru oxigen; în timpul oxigenării, atunci 2, 3-bisfosfogliceratul este expulzat.
În eritrocite există un aparat special care convertește 1, 3-bisfosfogliceratul (produs de metabolism) în 2, 3-bisfosfoglicerat astfel încât acesta să atingă o concentrație de 4-5 mM și, prin urmare, hemoglobina să poată schimba oxigen în țesuturi.
Hemoglobina care vine la un țesut este în stare eliberată (legată de oxigen), dar aproape de țesut, este carboxilată și trece într-o stare tensionată: proteina în această stare are tendința să se lege cu oxigen, comparativ cu starea eliberată, prin urmare hemoglobina eliberează oxigen în țesut; în plus, prin reacția dintre apă și dioxidul de carbon, există o producție de ioni de H +, apoi oxigen suplimentar pentru efectul bohr.
Dioxidul de carbon difuzează în eritrocite care trec prin membrana plasmatică; deoarece eritrocitele reprezintă aproximativ 40% din sânge, ar trebui să ne așteptăm ca numai 40% din dioxidul de carbon care se răspândește din țesuturi să intre în ele, de fapt, 90% din dioxidul de carbon intră în eritrocite deoarece conțin o enzimă care convertește dioxidul de carbon în acidul carbonic, rezultă că concentrația staționară de dioxid de carbon din eritrocite este scăzută și, prin urmare, viteza de intrare este ridicată.

Un alt fenomen care apare atunci când un eritrocit atinge un țesut este următorul: prin gradient, HCO3- (derivatul dioxidului de carbon) iese din eritrocite și, pentru a echilibra producția unei încărcări negative, avem care cauzează o creștere a presiunii osmotice: pentru a compensa această variație, este de asemenea introdusă apă care provoacă o umflare a eritrocitelor (efectul HAMBURGER). Fenomenul opus apare atunci când o eritrocită ajunge în alveolele pulmonare: există o deflație a eritrocitelor (efectul HALDANE). Prin urmare, eritrocitele venoase (direcționate către plămâni) sunt mai rotunde decât cele arteriale.


